
The Phytochrome System – Why Use Far-Red?
ANO004 // JOHANN WALDHERR // DR. RICHARD BLAKEY
1 Introduction
The light requirement of plants is now known to be far more complex than originally thought leading to the development of numerous LED technologies that produce a variety of different light spectra, both monochromatic and polychromatic. The worthwhile inclusion of some wavelengths in light recipes are as yet experimental, but one area of the spectrum that cannot be ignored is the far-red region. Far-red encompasses wavelengths 700 – 800 nm, a region of light that is on the edge of visibility in humans. However, these wavelengths have been proven to result in faster growth, increased biomass and better sensory characteristics (e.g. smell, taste, texture, color). But why are wavelengths that aren’t used in photosynthesis so influential on plant development? Unlike people and animals, plants are unable to move. Their sessile existence means that without any outside influence, plants will grow and live in the same place for all of their existence. This may seem like a simple observation but the consequence for plants is that they must be able to tolerate and survive when their immediate surroundings change to less favorable conditions. Responses to limited resources such as water, nutrients and light, in addition to circadian and circannual cycles are essential for plant survival. These responses can be manipulated to achieve favorable growth characteristics. This application note describes why these survival techniques evolved and why far-red wavelengths are essential for plant luminaires. For an introduction to the use of LEDs in horticultural applications, please refer to ANO002 LEDs - The Future of Horticultural Lighting.
2 Photoreceptors and the Phytochrome System
Light is primarily needed for photosynthesis, the main energy conversion mechanism of a plant and the main development factor that is mainly driven by red and blue light via chlorophyll in photosystems II and I. Three factors are important here:
- Light intensity - which is the amount of photons that the plant can use.
- Photoperiodism - which reflects the duration of the exposure.
- Light quality - which corresponds to the wavelengths of light plants are exposed.
But light also influences a number of other plant processes. Each process can be linked to a photoreceptor that reacts to a specific range of wavelengths. Cryptochromes sense blue/UVA light and are responsible for phototropism and photomorphogenesis, whereas photoreceptors called phytochromes detect far-red light (Figure 1). Phytochromes are unlike cryptochrome blue light receptors, as the phytochrome system is intrinsically reliant on the interplay between two wavelengths. The system consists of two forms of phytochromes that differ in their absorption wavelengths.[1] Pr (Phytochrome red) has an absorption maximum at 660 nm and Pfr (Phytochrome farred) an absorption maximum at 730 nm. However, interestingly Pr and Pfr can reversibly interconvert their molecular structure depending on the ratio of red and far-red wavelengths (Figure 2).
Figure 1: Typical absorbance spectra of the principle pigments of plants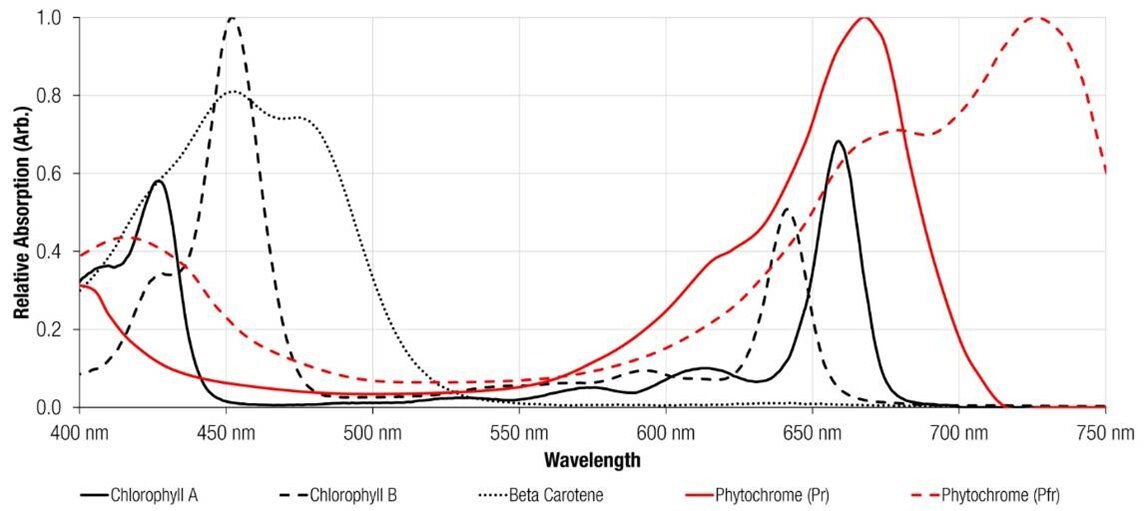
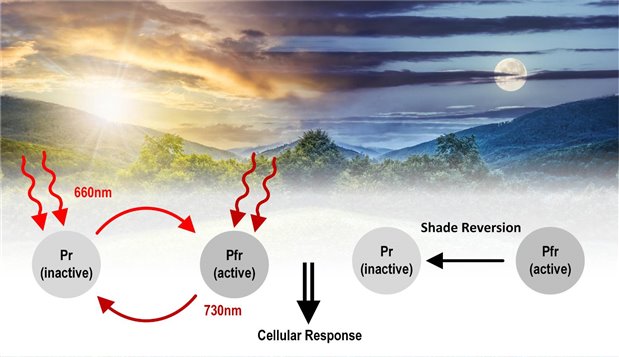
Figure 2: Functionality of phytochrome system
Pfr photoreceptors are considered the active form being converted from the Pr form in the presence of 660 nm red light. Pfr is physiologically active, initiating biological responses but is unstable. This means in cases of diminished or absent 660 nm light, it will revert to the Pr form. The Pfr form is also converted to the inactive Pr form in the presence of 730 nm far red light. Therefore, the ratio (light quality) in addition to the exposure time (photoperiodism) and total amount (light intensity) of red and far-red wavelengths a plant is exposed to, effects the phytochrome system. Different ratios of red/far-red wavelengths can trigger biological pathways that can significantly affect the desirable characteristics of plants.
3 Morphological responses to red and farred light
The phytochrome system controls a variety of molecular processes, which effect a number of morphological changes that allow plants to adapt to its light environment. These responses to red and farred light evolved as responses to the environment and in some cases are survival methods. Namely, the shade avoidance system for cases where a plant is not receiving enough light. This can either be a result of a solid object (obscuring all light) or other plants (transmitting some wavelengths of light). Another corresponds to changes in daily and seasonal light, which correspond to variations in temperature and humidity and the response of the plant to these conditions.
3.1. Light quality and the shade avoidance syndrome
In some ecosystems, plants can grow in extremely high densities. In this environment, plants compete for a limited amount of light, which drives photosynthesis. This can be between plants growing at the same time or growing in the shade of a taller plant. In order to compete and survive in this environment, plants sense the amount
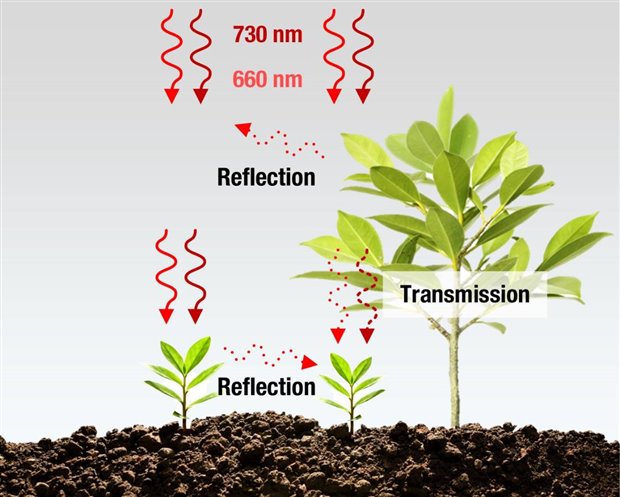
Figure 3: The reflected and transmitted light paths of red and far-red wavelengths in plant ecosystems
of shade they are in using the phytochrome system. The red and far red light reaching a plant in a light competitive environment arrives via a number of routes. The majority of light from the far-red region is either reflected or transmitted by plant material, lowering the ratio of red/far-red reaching plants in the immediate surroundings. This can occur as reflection from surrounding plants or transmitted light through a mature canopy (Figure 3). This means that direct sunlight has a high proportion of red light while light transmitting or reflecting from leaves is red deficient while having a relatively higher proportion of far red light. The red to far-red ratio mediates the control of the phytochrome system and consequently the shade avoidance response in plants that are shade intolerant or sun loving. Numerous studies have shown these responses to include accelerated elongation of hypocotyls, internodes, and petioles, elevated leaf angles to the horizontal and reduced branching in an attempt to capture more sunlight and drive photosynthesis.[2] This behavior is to ensure the survival of the plant. Additionally, the shade avoidance response can trigger early flowering whereby the plant reduces growth and starts the reproduction phase in an attempt to ensure.[3] The practical applications of this depends on the desirable characteristics of the plant being grown. In lettuce, germination is inhibited by far red light,[4] but it is important to know that the germination response here depend on the last light treatment.[5] Supplementation with far red light during the growth stage results in increased shoot and root growth with higher shoot fresh weight, and leaf area.[6] Eucalyptus cuttings have greater rooting success under low red to far-red ratios.[7] Far-red light can increase yields of green beans [8] and promote taller tomato plants.[9] Interestingly, farred can also be used to inhibit stem growth in plants where it is undesirable to grow too tall, reducing or even eliminating the use of growth inhibitors. The phytochrome system also controls carbon allocation and the metabolic status in developing plants.[10]
3.2. Photoperiodism and circadian rhythm
Plants are also sensitive to the shift of wavelengths from red to blue light that occurs at sunrise and the opposite shift at sunset. What’s more, they can perceive the time these daily events happen, which will affect plant processes like flowering. Like most life forms, the natural rhythm of the world must be adapted to with periods of activity and periods of rest to limit energy expenditure to when it will be most effective and beneficial. For example, it is usually nonbeneficial for a plant to flower during the night or in winter when there are few or no pollinators active during these periods. As plants develop, there is a vegetative phase (growth) and a reproductive phase (flowering). While photosynthesis during the growth phase causes the plants, leaves and shoots to develop, almost no growth takes place in the flowering phase. Through the development of flowers, plants can be pollinated and produce fruits or seeds. The process of flower production is irreversible once triggered and so the timing of this phase is crucial.[11] The targeted influencing of the flower formation plays an enormously important role especially for ornamental plant and seed producers. The phytochrome system is the key system for identifying and reacting to these changes. Phytochrome Pr is produced by the plant during darkness and accumulates. The Pfr generated during the day by far-red light, slowly reverts to the inactive Pr form (half-life = 2.5 h) in a process called dark reversion. During the day, the Pr form is converted to Pfr, restoring the Pr/Pfr equilibrium. Therefore, during night periods the Pr/Pfr ratio is high while during daytime the ratio it is low. The effect of the phytochrome system’s response to these changes depends on the type of plant. Plants can be divided into long-day plants (LDP) and short-day plants (SDP). SDP require a dark phase to flower, which must not be interrupted by light. If this dark phase is not reached, the plant will not flower. Conversely, LTP require a light phase in order to flower (Figure 4).[11] The hyper-red LED emits 660 nm light in the red spectral range and the 730 nm far-red LED in the dark red spectral region. A plant exposed to more red light relative to blue light, with a low red/farred ratio will interpret this as the start of a new day, potentially triggering flowering. As the ratio increases, the plant knows the transition has been made to daytime. As the ratio begins to fall again, caused by the sun setting, the plant senses it is getting dark and so metabolic and morphological changes begin for scotophile (night loving) processes. The length of the night can be extremely influential on the morphology of the plant.[12] The practical applications of this are numerous depending on the type of plant being cultivated. Ornamental plant growers can determine through the targeted manipulation of the phytochrome system when products are ready for market. This is extremely advantageous when you consider the annual demand cycles for fresh ornamental flowers. Days such as Valentine’s Day and Mother’s Day when demand is highest are early in the year, outside of the typical flowering time for most ornamental plants. But additionally, the plant can also be forced to flower several times a year. Under artificial light the growth/flower cycles are not tied to the seasons. Therefore, there is the opportunity to bring the plant to flower in the low light conditions of winter.

Figure 4: The effect of red/far-red photoperiodism on the flowering of plants
Likewise, unfavorable blooming times can be avoided during the summer months. For Example the Model plant Arabidopsis thaliana is a long-day plant and flowers earlier under long-day conditions than under short day conditions.[5] Chrysanthemum is a qualitative short day plant. Uniform flowering can be achieved with a photoperiod of 13.5 h or less. When the photoperiod is longer, no flowering occurs.[13] In cut chrysanthemum cultivation, the grower also uses long day conditions for around 2-3 weeks before switching to short day conditions, because of the required stem length specification.[12] Interestingly, when the night period is interrupted with hyper red light, the flowering in Chrysanthemum can be inhibited. This is also the case in Perilla plants [14] and wheat.[15] Therefore, the duration of the night period can be more important than the duration of the light period. The flowering of Kalanchoe, a short day plant, is mainly controlled by red light.[16] With this information, growers can develop their own optimized light-recipe for cultivated plants.
4 Summary
Phytochromes are photoreceptors and are one of the most important ways of plants to perceive their environment. They are responsible for detecting red and far-red light that initiate and control a variety of responses in the different development stages of plants making. The functional principle of phytochrome Pr (660 nm) and Pfr (730 nm) can be used to control many photomorphological processes in a plant. The exact affect depends on the species and cultivar. With the help of the horticultural expertise, it can lead to an increase in desirable quality parameters, making the use of both red and far red light in plant luminaires essential. It is advantageous for growers to be able to control the 730 nm LEDs separately and to adjust them according to their needs or their set quality parameters. Depending on the objective, it makes sense for the LEDs with the wavelength 730 nm to be separately dimmable in a single channel.
For this reason, we have developed specialized horticulture LEDs with wavelengths of 660 nm (Hyper Red) and 730 nm (Far Red) found in our WL-SMDC Horticulture series. These products have been developed to match the absorption peak of the two phytochrome forms (Figure 5).
Figure 5: The absorption spectra of phytochromes Pr and Pfr and the light output of the WL-SMDC hyper-red (150353HS74500) and far-red (150353FS74500) Ceramic LEDs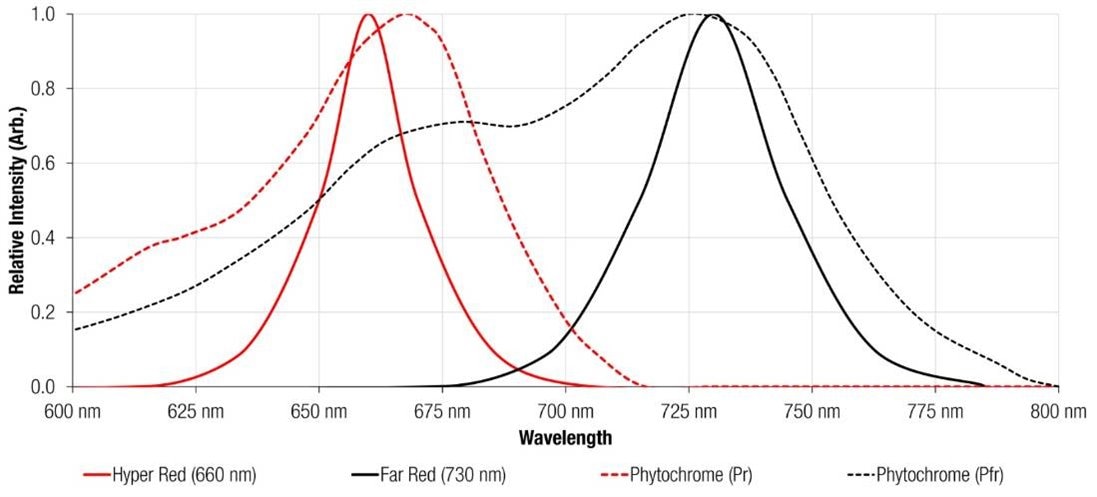
Figure 6: Würth Elektronik WL-SMDC SMD Mono-color Ceramic LED Waterclear
A. Appendix
A.1. Bill of Material
| Index | Description | Size | Value | WE Order Code | Farnell Order Code |
|---|---|---|---|---|---|
| WL-SMDC | 3535 | Hyper-red λ = 660 | 150353HS74500 | 2900781 | |
| WL-SMDC | 3535 | Far-red λ = 730 | 150353FS74500 | 2900780 |
A.2. Bibliography
[1] J. Li, G. Li, H. Wang, and X. Wang Deng, “Phytochrome signaling mechanisms.,” Arab. B. , vol. 9, p. e0148, 2011.
[2] R. Pierik and M. de Wit, “Shade avoidance: phytochrome signalling and other aboveground neighbour detection cues,” J. Exp. Bot. , vol. 65, no. 11, pp. 2815– 2824, Jun. 2013.
[3] K. J. Halliday, M. G. Salter, E. Thingnaes, and G. C. Whitelam, “Phytochrome control of flowering is temperature sensitive and correlates with expression of the floral integrator FT.,” Plant J., vol. 33, no. 5, pp. 875–885, Mar. 2003.
[4] S. Contreras, M. A. Bennett, J. D. Metzger, D. Tay, and H. Nerson, “Red to Far-red Ratio During Seed Development Affects Lettuce Seed Germinability and Longevity,” HortScience , vol. 44, no. 1, pp. 130–134, Feb. 2009.
[5] H. Wang and H. Wang, “Phytochrome Signaling: Time to Tighten up the Loose Ends,” Mol. Plant, vol. 8, no. 4, pp. 540–551, Apr. 2018.
[6] M.-J. Lee, K.-H. Son, and M.-M. Oh, “Increase in biomass and bioactive compounds in lettuce under various ratios of red to far-red LED light supplemented with blue LED light,” Hortic. Environ. Biotechnol. , vol. 57, no. 2, pp. 139–147, Apr. 2016.
[7] S. P. Hoad and R. R. B. Leakey, “Effects of pre-severance light quality on the vegetative propagation of Eucalyptus grandis W Hill ex Maiden - Cutting morphology, gas exchange and carbohydrate status during rooting,” TREES-STRUCTURE Funct., vol. 10, no. 5, pp. 317–324, Jun. 1996.
[8] P. A. Davis and C. Burns, “Photobiology in protected horticulture,” Food Energy Secur. , vol. 5, no. 4, pp. 223–238, 2016. [9] C. Kubota, P. Chia, Z. Yang, and Q. Li, “Applications of far-red light emitting diodes in plant production under controlled environments,” in International Symposium on Advanced Technologies and Management Towards Sustainable Greenhouse Ecosystems: Greensys2011 952 , 2011, pp. 59–66.
[10] Z.-C. Yang, C. Kubota, P.-L. Chia, and M. Kacira, “Effect of end-of-day far-red light from a movable LED fixture on squash rootstock hypocotyl elongation,” Sci. Hortic. (Amsterdam). , vol. 136, pp. 81–86, 2012.
[11] F. Tooke, M. Ordidge, T. Chiurugwi, and N. Battey, “Mechanisms and function of flower and inflorescence reversion,” J. Exp. Bot., vol. 56, no. 420, pp. 2587– 2599, 2005.
[12] Y. H. Song, J. S. Shim, H. A. Kinmonth-Schultz, and T. Imaizumi, “Photoperiodic Flowering: Time Measurement Mechanisms in Leaves,” Annu. Rev. Plant Biol., vol. 66, pp. 441–464, Apr. 2015.
[13] S. Kumar M.C. Singh and D. Sharma, Effect of Photosynthetically Active Radiation (PAR) from LEDs on Growth and Development of Chrysanthemum morifolium Ramat. cv. Zembla, vol. 6. 2017.
[14] K.-Y. Park, Flowering Response to Light Intensity and Night Interruption in Perilla , vol. 40. 1995.
[15] S. Pearce, L. M. Shaw, H. Lin, J. D. Cotter, C. Li, and J. Dubcovsky, “Night-break experiments shed light on the Photoperiod 1-mediated flowering,” Plant Physiol. , Jan. 2017.
[16] W. Amaki and M. Kunii, “Effects of light quality on the flowering responses in Kalanchoe blossfeldiana,” in Acta Horticulturae , 2015, no. 1107, pp. 279–284.
IMPORTANT NOTICE
The Application Note is based on our knowledge and experience of typical requirements concerning these areas. It serves as general guidance and should not be construed as a commitment for the suitability for customer applications by Würth Elektronik eiSos GmbH & Co. KG. The information in the Application Note is subject to change without notice. This document and parts thereof must not be reproduced or copied without written permission, and contents thereof must not be imparted to a third party nor be used for any unauthorized purpose.
Würth Elektronik eiSos GmbH & Co. KG and its subsidiaries and affiliates (WE) are not liable for application assistance of any kind. Customers may use WE’s assistance and product recommendations for their applications and design. The responsibility for the applicability and use of WE Products in a particular customer design is always solely within the authority of the customer. Due to this fact it is up to the customer to evaluate and investigate, where appropriate, and decide whether the device with the specific product characteristics described in the product specification is valid and suitable for the respective customer application or not. The technical specifications are stated in the current data sheet of the products. Therefore the customers shall use the data sheets and are cautioned to verify that data sheets are current. The current data sheets can be downloaded at www.we-online.com. Customers shall strictly observe any product-specific notes, cautions and warnings. WE reserves the right to make corrections, modifications, enhancements, improvements, and other changes to its products and services.
WE DOES NOT WARRANT OR REPRESENT THAT ANY LICENSE, EITHER EXPRESS OR IMPLIED, IS GRANTED UNDER ANY PATENT RIGHT, COPYRIGHT, MASK WORK RIGHT, OR OTHER INTELLECTUAL PROPERTY RIGHT RELATING TO ANY COMBINATION, MACHINE, OR PROCESS IN WHICH WE PRODUCTS OR SERVICES ARE USED. INFORMATION PUBLISHED BY WE REGARDING THIRD-PARTY PRODUCTS OR SERVICES DOES NOT CONSTITUTE A LICENSE FROM WE TO USE SUCH PRODUCTS OR SERVICES OR A WARRANTY OR ENDORSEMENT THEREOF.
WE products are not authorized for use in safety-critical applications, or where a failure of the product is reasonably expected to cause severe personal injury or death. Moreover, WE products are neither designed nor intended for use in areas such
as military, aerospace, aviation, nuclear control, submarine, transportation (automotive control, train control, ship control), transportation signal, disaster prevention, medical, public information network etc. Customers shall inform WE about the intent of such usage before design-in stage. In certain customer applications requiring a very high level of safety and in which the malfunction or failure of an electronic component could endanger human life or health, customers must ensure that they have all necessary expertise in the safety and regulatory ramifications of their applications. Customers acknowledge and agree that they are solely responsible for all legal, regulatory and safety-related requirements concerning their products and any use of WE products in such safety-critical applications, notwithstanding any applications-related information or support that may be provided by WE.
CUSTOMERS SHALL INDEMNIFY WE AGAINST ANY DAMAGES ARISING OUT OF THE USE OF WE PRODUCTS IN SUCH SAFETYCRITICAL APPLICATIONS.
USEFUL LINKS
Application Notes
REDEXPERT Design Platform
Toolbox
Product Catalog
DIRECT LINKS
ANO004: The Phytochrome system – Why use far-red?
CONTACT INFORMATION
Tel. +49 7942 945 - 0
Würth Elektronik eiSos GmbH & Co. KG
Max-Eyth-Str. 1 / 74638 Waldenburg / Germany
Top Comments